Beyond Darwin: reactive heredity, burst-drift dynamics and eco‑evolutionary control of cancer
Laurine Lagache, Julien Salzet, Isabelle Fournier, Michel Salzet
Molecular Cancer
https://doi.org/10.1186/s12943-026-02683-w
Abstract
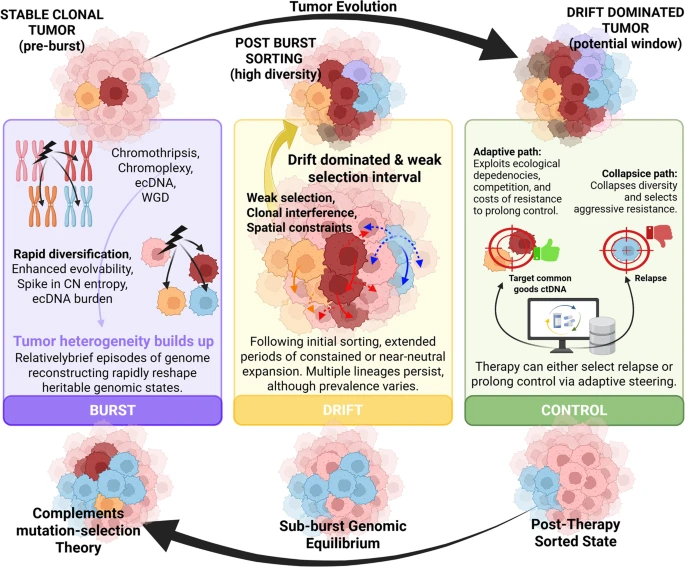
Darwinian clonal evolution provides a necessary framework for understanding tumour progression, yet emerging evidence shows that some cancers depart from a strictly gradualist tempo through episodic “macroevolutionary” leaps. We propose a unifying Burst-Drift-Control (BDC) model in which short-lived genomic reconfigurations (“bursts”) that transiently alter heritable genomic states (“reactive heredity”) precede longer intervals of constrained expansion, creating time-bounded vulnerabilities that clinicians may strategically steer (“control”). In this usage, the Drift component refers specifically to a drift-dominated or weak-selection interval after any initial post-burst sorting, not to the selective sweep itself. In the Burst phase, single catastrophic events, such as extrachromosomal DNA amplifications, chromothripsis, chromoplexy or whole-genome doubling, can generate major shifts in genotype and phenotype. These bursts produce new genetic configurations and can increase evolvability, for example when ecDNA circles permit oncogene dose tuning and non-Mendelian inheritance. During the drift-dominated interval, tumours may enter longer periods of constrained or near-neutral expansion in which spatial structure, clonal interference and weak or unresolved selection allow multiple lineages to persist, although the prevalence of such dynamics varies substantially by tumour type and treatment context. BDC does not assume that selective advantage is higher immediately after a burst than at later time points; later therapy or microenvironmental change can also impose strong selection. Finally, in the Control phase, if therapy is timed to exploit ecological dependencies and the fitness costs of resistance, cancer growth may be restrained as a stable, polyclonal “oligopoly” rather than progressing rapidly to an aggressive monoclonal takeover. We discuss supporting evidence, counterexamples and practical limitations, and argue that the main value of BDC lies in linking punctuated genome change to testable windows for evolutionary steering.